¿Qué son los protozoos?
El desciframiento de las raíces griegas lleva a definir los protozoos como "primeros" (proto) "animales" (zoa). Aunque los estudios filogenéticos moleculares indican que los protozoos se encuentran entre los primeros eucariotas ramificados (véase el árbol filogenético), tal definición no aporta mucha información descriptiva. Los protozoos no son fáciles de definir porque son muy diversos y a menudo sólo están lejanamente relacionados entre sí. Debido a la extrema diversidad de los protozoos, la única característica común a todos ellos es que son microorganismos eucariotas unicelulares. Los protozoos poseen orgánulos eucariotas típicos y, en general, presentan las características típicas de otras células eucariotas. Por ejemplo, en todas las especies de protozoos existe un núcleo unido a una membrana que contiene los cromosomas. Sin embargo, en muchas especies de protozoos algunos de los orgánulos pueden estar ausentes o ser morfológica o funcionalmente diferentes de los que se encuentran en otros eucariotas. Además, muchos de los protozoos tienen orgánulos que son exclusivos de un grupo concreto de protozoos.
Las preguntas que uno puede hacerse para definir a los protozoos son:
- ¿Qué aspecto tienen?
- ¿Qué tamaño tienen?
- ¿Dónde viven?
- ¿Cómo se desplazan?
- ¿Cómo se reproducen?
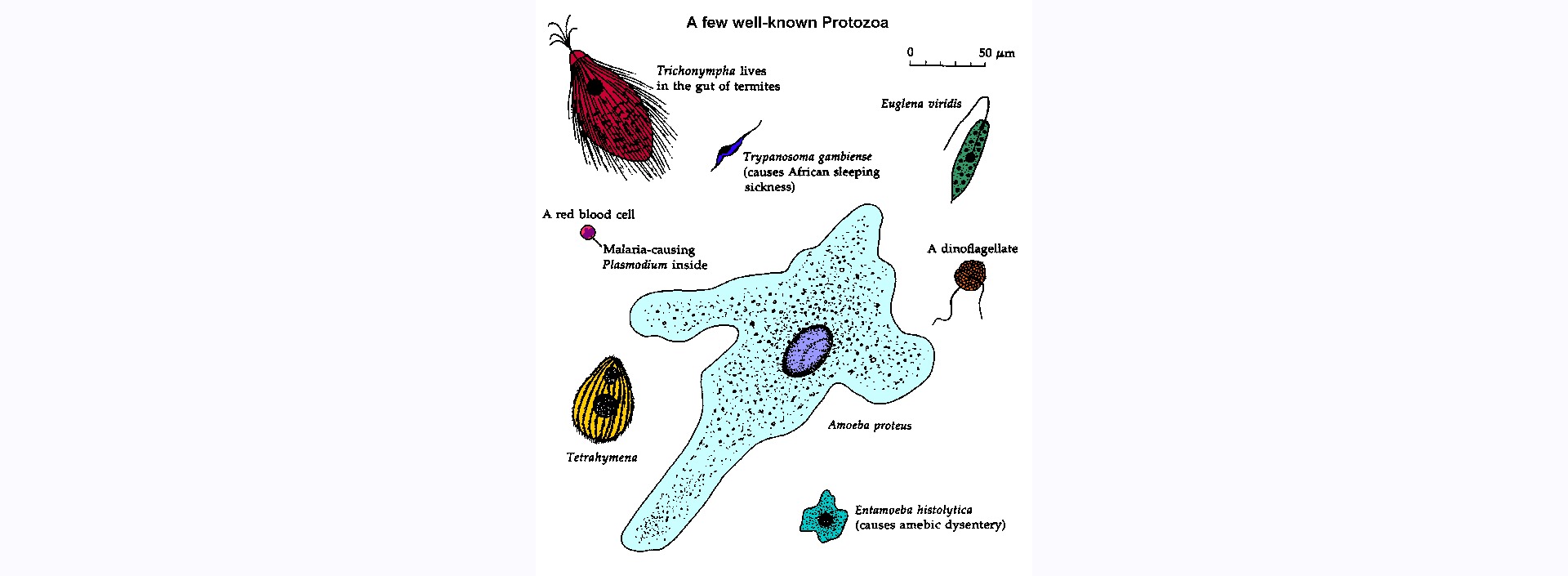

Los protozoos se encuentran en ambientes húmedos prácticamente en todas partes. Como grupo, los protozoos son extremadamente adaptables. Sin embargo, las especies individuales suelen tener nichos específicos. Como todos los demás organismos, los protozoos deben ser capaces de adquirir y metabolizar nutrientes de su entorno (es decir, heterótrofos). Muchos protozoos simplemente absorben solutos (osmotrofia) de su medio, mientras que otros son carroñeros que ingieren material sólido (fagotrofia). Los protozoos depredadores cazan activamente o emboscan pasivamente a otros organismos (normalmente bacterias u otros protozoos). Algunos protozoos son fotosintéticos y pueden captar la energía del sol y convertirla en energía química utilizable (es decir, autótrofos o fotótrofos). Muchos protozoos no se limitan a un único mecanismo de alimentación y pueden utilizar combinaciones de los anteriores (es decir, mixótrofos).
Los protozoos también pueden considerarse de vida libre o simbióticos. Por lo general, los organismos de vida libre se encuentran en el suelo o en medios acuosos, mientras que los simbiontes viven en estrecha asociación con otro organismo. La simbiosis implica una dependencia fisiológica de un organismo respecto a otro y no sólo una estrecha asociación física entre dos organismos. Por lo general, esta dependencia se da en forma de nutrición. Se pueden distinguir diferentes formas de simbiosis que reflejan la naturaleza de la asociación entre los dos organismos (recuadro).

Las primeras observaciones de los protozoos destacaron su motilidad. Esta motilidad dio lugar a su clasificación como "animales", que se distinguían de las "plantas" no móviles. Sin embargo, la motilidad no es una característica universal de los protozoos y los distintos protozoos utilizan diferentes mecanismos para su movimiento (Tabla). De hecho, los protozoos se clasificaron inicialmente basándose en parte en su mecanismo de motilidad (véase Taxonomía).
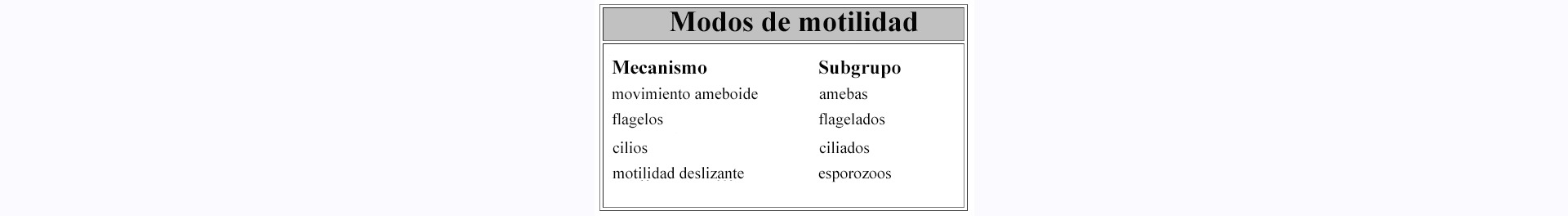
A diferencia de los flagelados y los ciliados, las amebas son protozoos que se arrastran a lo largo de un sustrato sólido de una forma conocida como "movimiento ameboide". La ameba proyecta un pseudopodio, o pie falso, desde el cuerpo celular. El pseudopodio se adhiere al sustrato y tira del resto del cuerpo celular hacia delante. La fuerza implicada en este movimiento la genera otro sistema citoesquelético, compuesto por actina y miosina. La actina forma largos filamentos, también conocidos como microfilamentos, y la miosina es una proteína motora que se desplaza a lo largo de los microfilamentos de forma dependiente del ATP. La contracción muscular es otro ejemplo de generación de fuerza a través de elementos del citoesqueleto actina-miosina. En un sentido mecanicista, la fagocitosis es una forma de movimiento ameboide que también implica a los microfilamentos. En este caso, los pseudópodos se extienden para rodear la partícula que se está ingiriendo. La fusión de los pseudópodos con el cuerpo celular da lugar a la internalización de la partícula dentro de una vacuola.
Los apicomplexa también se arrastran por un sustrato, pero por un mecanismo diferente al de las amebas. El mecanismo de esta llamada "motilidad deslizante" está empezando a comprenderse y probablemente implique sistemas citoesqueléticos basados tanto en microfilamentos como en microtúbulos. Los Apicomplexa también presentan formas intracelulares y la invasión de la célula huésped también implica esta motilidad deslizante. (Véase también la discusión sobre la invasión de la célula huésped por el parásito de la malaria).
La motilidad celular implica la generación de fuerza a través de los elementos del citoesqueleto basados en microtúbulos o los elementos del citoesqueleto basados en microfilamentos. Esto es válido tanto para los protozoos como para otros eucariotas. La implicación de los microtúbulos y los microfilamentos tanto en la forma como en el movimiento celular hace que estas estructuras subcelulares sean más análogas al sistema musculoesquelético.
Reproducción

Muchos protozoos presentan reproducción sexual además de las formas asexuales de reproducción. Esta reproducción sexual puede implicar la producción y fusión de gametos en procesos similares a los de los organismos superiores. Los Ciliophora experimentan una conjugación en la que tipos de apareamiento opuestos se emparejan e intercambian directamente material genético (es decir, ADN). A veces, la reproducción sexual es un paso obligatorio en el ciclo vital, mientras que en otros casos el organismo puede reproducirse asexualmente con una ronda ocasional de reproducción sexual.
En resumen, los protozoos son microorganismos eucariotas unicelulares. Sin embargo, la gran diversidad en cuanto a morfología, tamaño y formas de vida que presentan los protozoos dificulta la elaboración de una definición más precisa. Su larga historia evolutiva (véase el árbol filogenético) explica gran parte de esta diversidad. Sin embargo, los protozoos presentan características comunes a todos los eucariotas. (Enlace a una serie de presentaciones en powerpoint sobre la biología de las células eucariotas).

Taxonomía
La taxonomía, o sistemática, es la ciencia que se ocupa de nombrar y clasificar los organismos. Además de asignar clasificaciones taxonómicas jerárquicas, la sistemática también intenta situar a los organismos en grupos que reflejen relaciones evolutivas o filogenias. Sin embargo, los criterios taxonómicos suelen ser arbitrarios y la taxonomía cambia constantemente para reflejar nuevos descubrimientos e interpretaciones. Además, en la sistemática de los microorganismos patógenos se utilizan con frecuencia características utilitarias, como el tipo de enfermedad, el rango de hospedadores y la distribución geográfica. Esto es especialmente cierto en el caso de la taxonomía de los protozoos. Además, existe cierto debate sobre la filosofía general en la clasificación de los protozoos y se desconocen las relaciones entre muchos grupos de protozoos (véase D.J. Patterson en Am. Nat. 154, S96-124). La clasificación de los protozoos se complica aún más por la definición original de los protozoos como heterótrofos unicelulares. Ahora se reconoce que los protozoos (o protistas) pueden utilizar múltiples estrategias nutricionales y no pueden considerarse simplemente como vegetales (autótrofos) o animales (heterótrofos). Por tanto, el término protozoo no puede considerarse un verdadero grupo taxonómico. A pesar de que la palabra protozoo per se ya no es un nombre taxonómico apropiado, sigue siendo un término útil y funcional.
Históricamente, los protozoos se dividían en cuatro grandes grupos: las amebas, los flagelados, los ciliados y los esporozoos. Los rasgos distintivos entre los grupos se basaban en la motilidad (es decir, ameboides, flagelos, cilios). Los esporozoos eran un grupo heterogéneo que producía esporas durante una fase de su ciclo vital y presentaba una motilidad "deslizante". Sin embargo, este esquema de clasificación es bastante arbitrario y no refleja necesariamente las verdaderas relaciones evolutivas entre los organismos. Uno de los problemas de utilizar la motilidad como criterio taxonómico es que muchos protozoos utilizan diferentes tipos de motilidad durante las distintas fases de sus ciclos vitales. Por ejemplo, Naegleria existe en forma de ameba cuando el alimento es abundante y se transforma en flagelado cuando falta. En general, las amebas constituyen un grupo heterogéneo y todas derivan probablemente de los flagelados. De estos cuatro grupos de protozoos originales, sólo los ciliados siguen considerándose un taxón válido.
A partir de la década de 1960 se utilizó el microscopio electrónico para identificar características ultraestructurales que pudieran servir de criterio para agrupar a los protozoos. En muchos casos, la morfología conduce a una clasificación que sitúa a los organismos en grupos monofiléticos. Monofilético significa que todos los organismos de ese grupo derivan probablemente de un antepasado común. Por ejemplo, muchos de los protozoos antes llamados esporozoos poseen estructuras subcelulares, conocidas colectivamente como orgánulos apicales, y ahora forman un grupo monofilético llamado apicomplexa. Sin embargo, las estructuras subcelulares y las vías metabólicas pueden perderse en algunos linajes y situar a sus descendientes puede resultar problemático.
Durante la década de 1980 y hasta la actualidad se están aplicando técnicas moleculares a la taxonomía. Mediante la comparación de secuencias de ADN o proteínas se pueden deducir posibles historias y relaciones evolutivas. Los datos de secuencias moleculares han confirmado filogenias basadas en otros criterios, han zanjado algunos debates y han dado lugar a algunas sorpresas. Por ejemplo, los datos moleculares confirman que los apicomplexa son monofiléticos y, además, indican que están emparentados con los ciliados y los dinoflagelados. Estos tres grupos se combinan ahora en un grupo monofilético mayor denominado alveolata. Esta relación ya se sospechaba y el nombre hace referencia a las estructuras morfológicas conocidas como sacos alveolares. Pero el uso de genes individuales puede ser un medio poco fiable para determinar las relaciones evolutivas, especialmente entre organismos emparentados a distancia, y los datos moleculares deben interpretarse con cautela.
Desde mediados de los años ochenta, la clasificación de los protistas ha sido objeto de controversias filosóficas. Por un lado, existen argumentos a favor de conservar elementos del esquema bütschliano basado en la motilidad debido a su familiaridad y simplicidad. Sin embargo, estos esquemas suelen entrar en conflicto con los datos filogenéticos. Además, las jerarquías y rangos de la sistemática linneana tradicional (es decir, filo, clase, orden, familia, género, especie) no siempre se ajustan bien a los microorganismos, y a menudo es difícil decidir qué nivel jerárquico es el más apropiado para un grupo de protozoos concreto.
Anteriormente, muchos esquemas taxonómicos han definido cinco reinos de la vida: procariotas (bacterias), protistas, plantas, hongos y animales. En estos esquemas, los protozoos forman parte de los protistas, junto con las algas unicelulares, las diatomeas, los oomicetos y los mohos limosos. Sin embargo, siempre ha habido insatisfacción con el grupo de los protistas. Esto se debe en parte a que los protistas se definen en parte por un criterio negativo. En otras palabras, los organismos que no encajan en los otros cuatro reinos se incluyen por defecto en los protistas. Además, algunos protistas están filogenéticamente más emparentados con los otros tres reinos eucariotas que con otros protistas, por lo que los protistas son claramente polifiléticos.
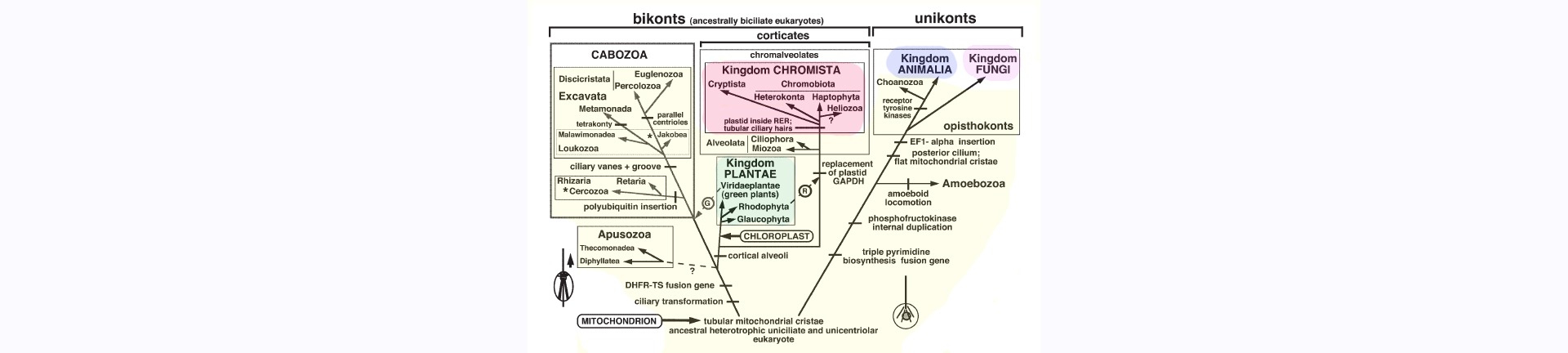
Cavalier-Smith propone cinco reinos eucariotas que consisten en el reino basal, y por tanto parafilético, Protozoa y cuatro reinos derivados: Animalia, Fungi, Plantae y Chromista (Figura). Algunos de los antiguos protistas se incluyen ahora en los reinos derivados con los que están más estrechamente relacionados, por lo que el reino Protozoa se convierte en monofilético. Sin embargo, los alveolados, que incluyen a muchos protozoos tradicionales como los apicomplexa, los ciliados y los dinoflagelados, forman un clado con los cromistas, pero se incluyen en el reino protozoo. Por tanto, los cromistas no son holofiléticos. Trasladar los alveolados a un nuevo reino llamado Chromalveolata resolvería este problema. Sin embargo, esto probablemente daría lugar a controversia y confusión, ya que muchos de los alveolados se han considerado durante mucho tiempo protozoos.
ENLACES
- ¿Qué son los protozoos? (Arriba)
- [Diversidad de los protozoos]
- Simbiosis
- Motilidad
- Reproducción
- Taxonomía
- Protozoología Inicio
- Biología Celular Eucariota
- Otros cursos y conferencias
- Wiser Inicio
Original article: www2.tulane.edu/~wiser/protozoology/notes/INTRO.html#symbiosis




